- 移动端


和元生物技术(上海)股份有限公司品牌商
14 年
手机商铺
- NaN
- 0.5
- 0.5
- 1.5
- 0.5

推荐产品
公司新闻/正文
Nat Commun : 武汉大学郑凌团队发现脂肪褐变新靶点
982 人阅读发布时间:2023-09-21 17:20
脂肪组织(WAT)是全身性能量代谢的关键器官, 脂肪组织功能紊乱与肥胖和二型糖尿病相关的代谢疾病密切相关。脂肪细胞是脂肪组织中的主要细胞类型, 可以被分为白色脂肪细胞, 米色脂肪细胞和棕色脂肪细胞, 在冷刺激下, 具有棕色脂肪产热表型的米色脂肪细胞会被诱导褐变。白细胞介素10 (IL10) 是免疫细胞分泌的一种抗炎细胞因子, 可以通过与在成熟脂肪细胞表达的受体IL10rα结合, 抑制产热功能。
许多表观遗传因子能够参与产热调控。然而, 作为染色质结构和动力学的关键调节因子, 组蛋白变异在脂肪组织中的作用尚不清楚。此前, macroH2A1.1在白色脂肪细胞分化中的关键作用已被报道, 突变的组蛋白H3.3在褐色前体脂肪细胞中的过表达会损害小鼠褐色脂肪细胞的发育, 这表明组蛋白变异可能参与了代谢稳态的调节。连接组蛋白突变体 (H1.2) 已经被报道在细胞凋亡, 自噬, DNA损伤, 肿瘤发生有关。然而, H1.2在新陈代谢中的功能,特别是在脂肪组织中的功能,仍然存在未知的。
2023年7月6日, Nature Communications刊登了武汉大学郑凌教授团队在脂肪组织领域的研究成果“Linker histone variant H1.2 is a brake on white adipose tissue browning”,研究团队通过RNA-sequence, ChIP-sequence,基因敲除, 过表达等多种手段结合, 发现了H1.2通过结合编码IL10受体的IL10rα启动子, 抑制产热基因的表达从而抑制米色脂肪组织产热。

01.脂肪组织产热闸口新靶点H1.2的发现
首先,研究者通过对十周龄雄性小鼠各组织的H1.2 进行蛋白含量测定, 发现H1.2在脂肪细胞中的表达量远高于其它组织 (图1a), 棕色脂肪组织 (BAT) 表达量最高, 腹股沟白色脂肪组织 (iWAT) 表达量其次, 附睾白色脂肪组织 (eWAT) 表达量最低 (图1b)。iWAT和BAT中的H1.2和解偶联蛋白1 (Ucp1) 的基因和蛋白表达量在低温(6℃, 22℃)下高于高温(22℃, 30℃)(图c-f), 同时, 其表达量随着脂肪分化程度和成熟程度逐渐升高 (图 g-l)。这些数据表明H1.2可能参与成熟米色和棕色脂肪细胞的适应性产热。
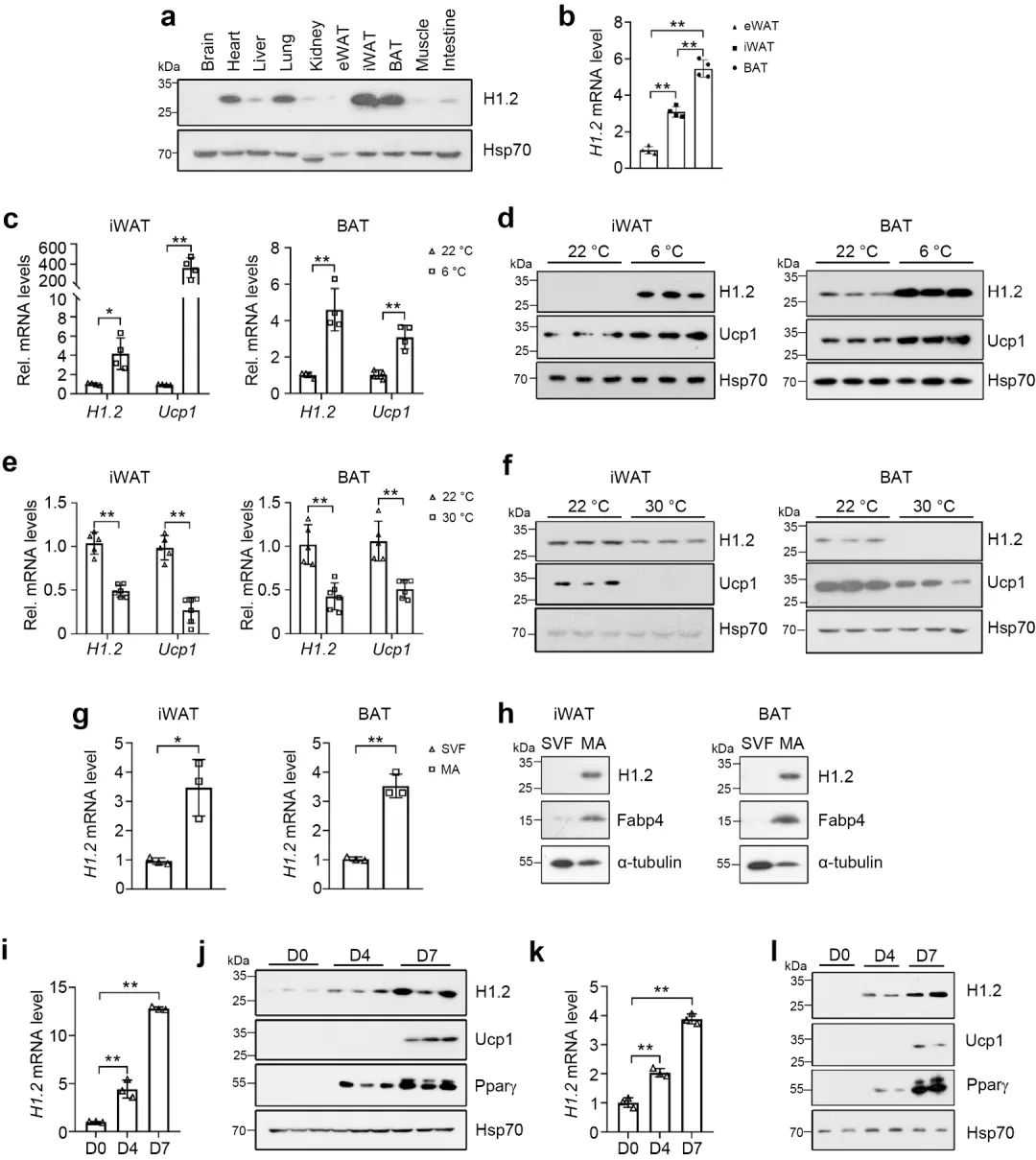
图1 米色/棕色脂肪细胞富含H1.2并能感知温度
02.脂肪细胞特异性H1.2缺失促进能量消耗
研究者进一步对小鼠脂肪组织中的H1.2进行基因敲除(H1.2AKO), 然后进行了RNA测序, 通过GO功能富集分析, 发现H1.2AKO小鼠中iWAT和BAT里面的差异基因主要参与PPARα信号通路, 与能量分解代谢密切相关 (图 2d-2e)。通过代谢笼实验, 发现H1.2AKO小鼠的氧气消耗量和能量消耗量高于WT小鼠, 白天作用更加显著 (图 2f-2g)。在H1.2AKO小鼠中, iWAT中的UCP1基因和蛋白表达量显著高于WT小鼠, 而BAT中的UCP1基因和蛋白表达量在H1.2AKO小鼠和WT小鼠中并没有显著区别 (图 2h-i)。这些数据表明H1.2缺失能够促进能量消耗, 并且在iWAT中发挥作用。
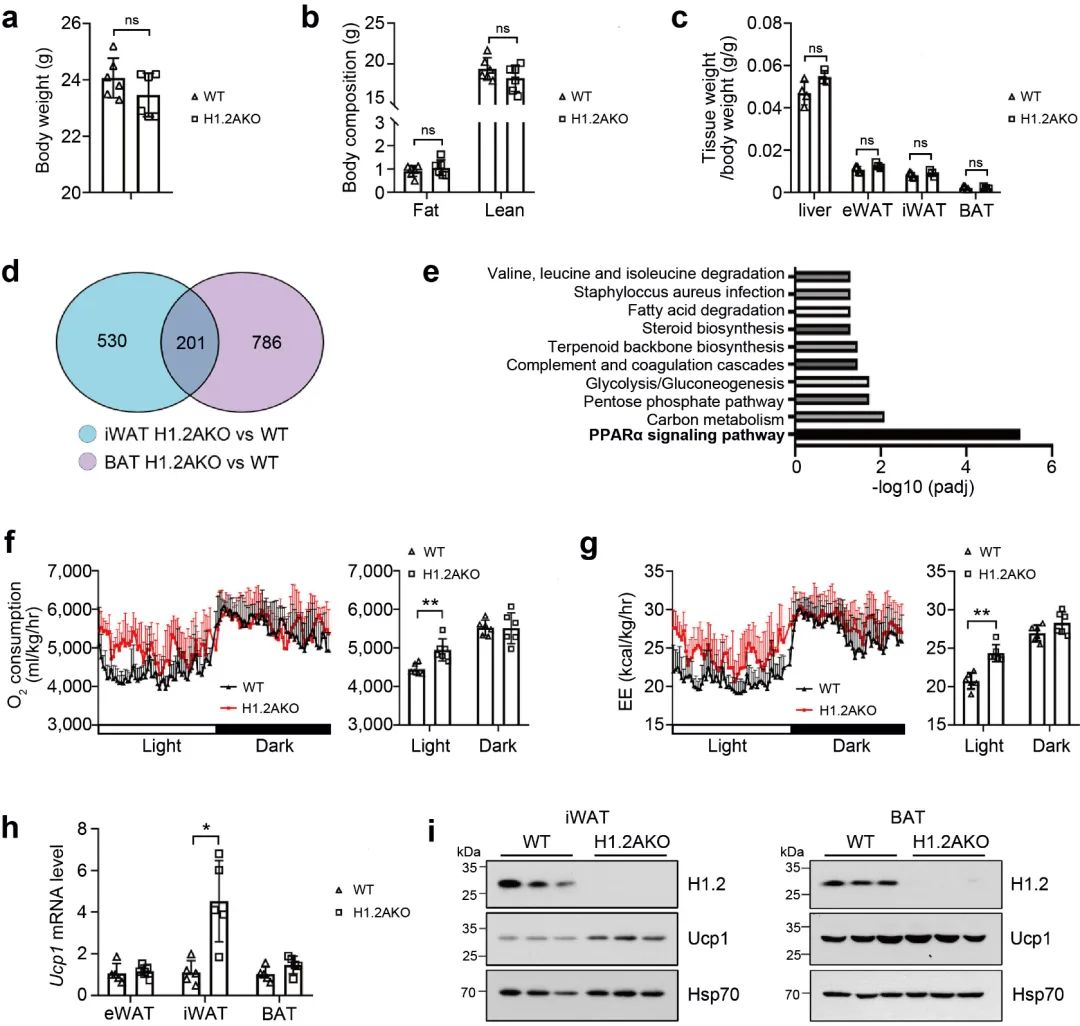
图 2 正常喂养的幼龄H1.2AKO小鼠能量消耗增加
03.脂肪细胞中的H1.2在冷刺激下诱导iWAT褐变
通过以上的研究, 可以推测出H1.2与iWAT产热有关。因此, 研究者将幼年H1.2AKO小鼠和WT小鼠在6℃下冷刺激3天, 发现H1.2AKO小鼠的体温升高, 氧气消耗增加, 能量消耗增加 (图 3a-c), 多腔的脂肪细胞增多, Ucp1表达量显著升高 (图 3d), 产热基因显著上调 (图 3e), 然而在冷暴露下的BAT中, H1.2AKO小鼠和和WT小鼠并没有显著区别。随后, 研究者对小鼠iWAT使用AAV作为载体进行H1.2过表达 (AAV-H1.2), 发现过表达H1.2组的小鼠iWAT中的Ucp1蛋白表达水平显著低于WT小鼠, 同样, 其体温显著降低, 表现出较低的耐寒性 (图 3f-h)。另外, 低温暴露后, H1.2 过表达小鼠的脂肪褐变受到抑制, Ucp1表达量降低, 产热基因的表达水平下降 (图 3i-k)。这些数据表明, 在冷暴露下, H1.2通过作用于iWAT而不是BAT, 作为适应性产热的负调节因子, 抑制iWAT褐变。
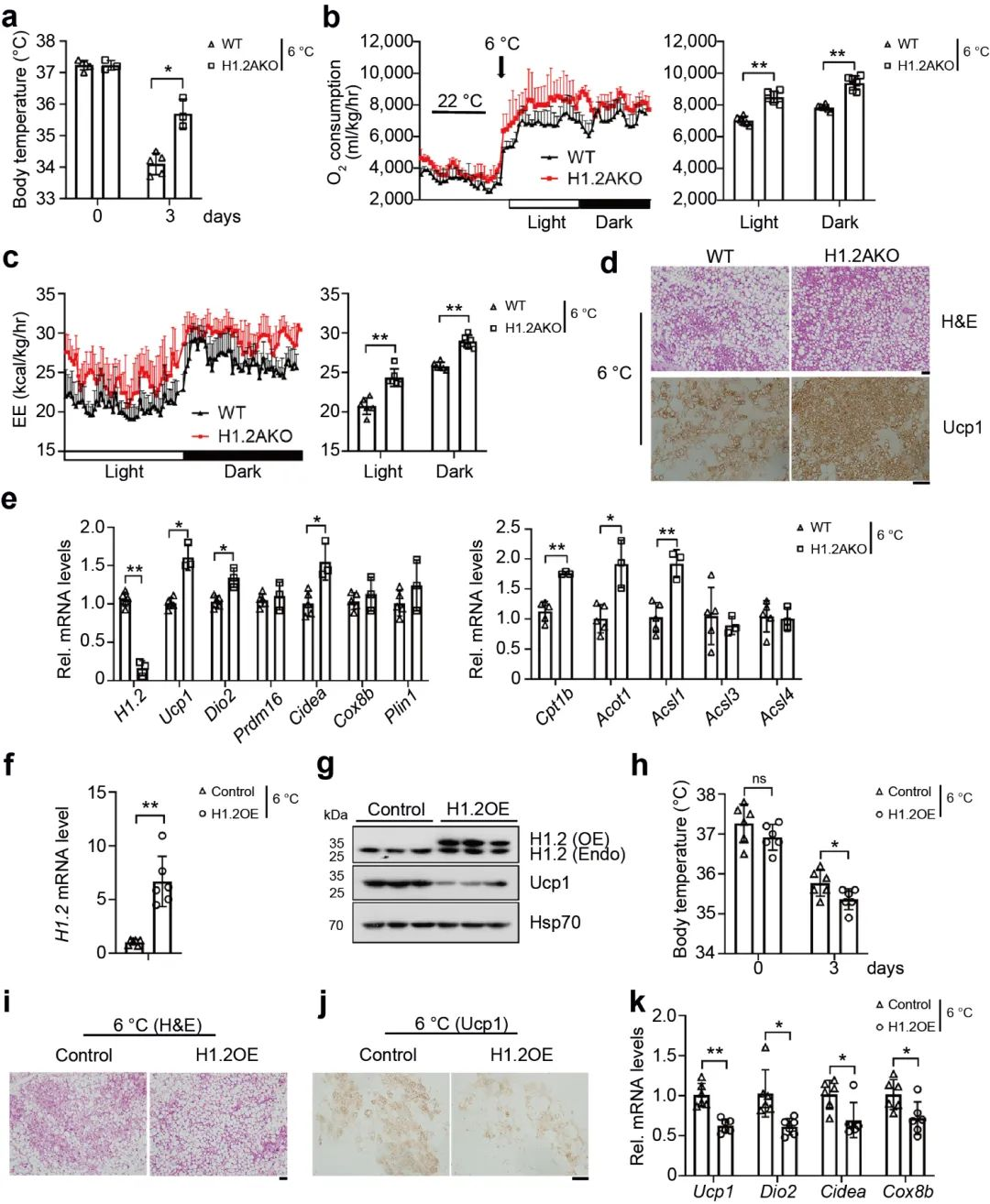
图 3 H1.2调节冷暴露下iWAT褐变
04.H1.2通过Il10rα调控米色脂肪细胞产热
基于上一系列的研究结果, 研究团队通过对H1.2AKO鼠的iWAT进行GO 富集分析, 发现显著改变的基因主要涉及到体液免疫应答的生物学过程 (图 4a)。随后, 通过qPCR检测了B细胞, T细胞, 和与这些细胞相关的趋化因子的几种生物标志物, 结果并没有出现显著性差异。然而, 在H1.2AKO小鼠的iWAT中, 发现IL10, IL10rα的表达量显著降低, 其IL10rβ的表达量并未受到显著影响 (图 4b-f)。另外, 我们发现H1.2AKO小鼠中的iWAT中的成熟的脂肪细胞中IL10rα的基因和蛋白的表达显著低于WT组, 冷刺激下效果更加明显, 而iWAT中的血管基质部分并没有显著差异 (图 4g-i)。
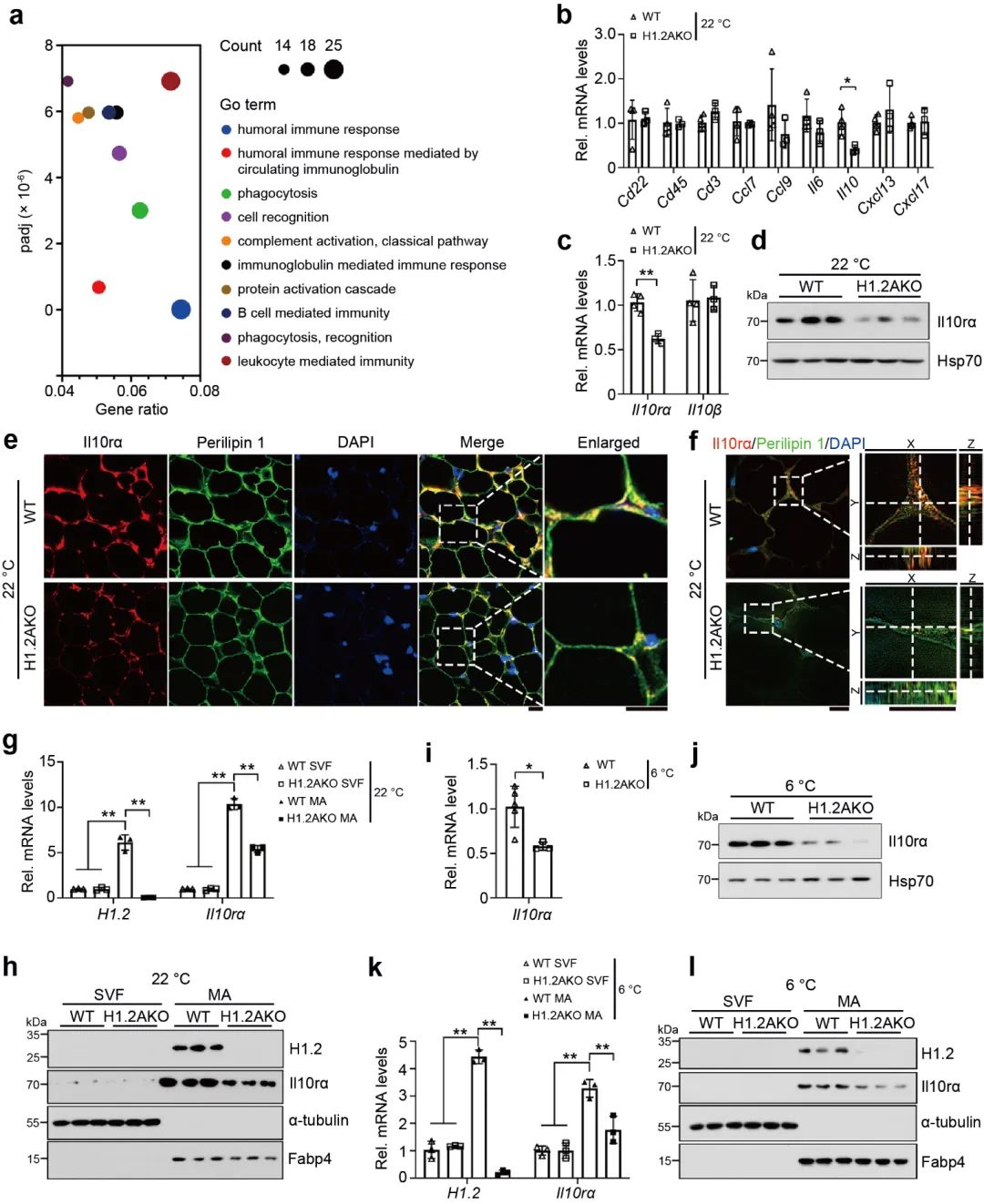
图4 IL10rα在H1.2AKO小鼠成熟脂肪细胞中下调
使用Cre重组酶 (Ad-Cre-H1.2) 处理H1.2 flox/flox小鼠的脂肪细胞, 发现H1.2缺乏的成熟的米色脂肪细胞或棕色脂肪细胞中一些产热基因, 以及脂质β-氧化基因显著升高 (图 5a-e)。同时, Ucp1蛋白水平显著上调, IL10rα水平显著降低 (图 5f)。在H1.2缺陷米色脂肪细胞, 对其进行异丙adrenaline处理, 同样发现IL10rα水平降低, Ucp1水平升高 (图 5g-h)。然而, 使用不同浓度的重组IL10蛋白来处理分化的米色脂肪细胞, 产热基因和蛋白的水平均下降, 在敲除H1.2后, IL10的这种产热抑制作用减弱 (图 5k-m)。这些数据表明, H1.2对于米色脂肪细胞的产热能力至关重要, H1.2 通过IL10rα 抑制产热。
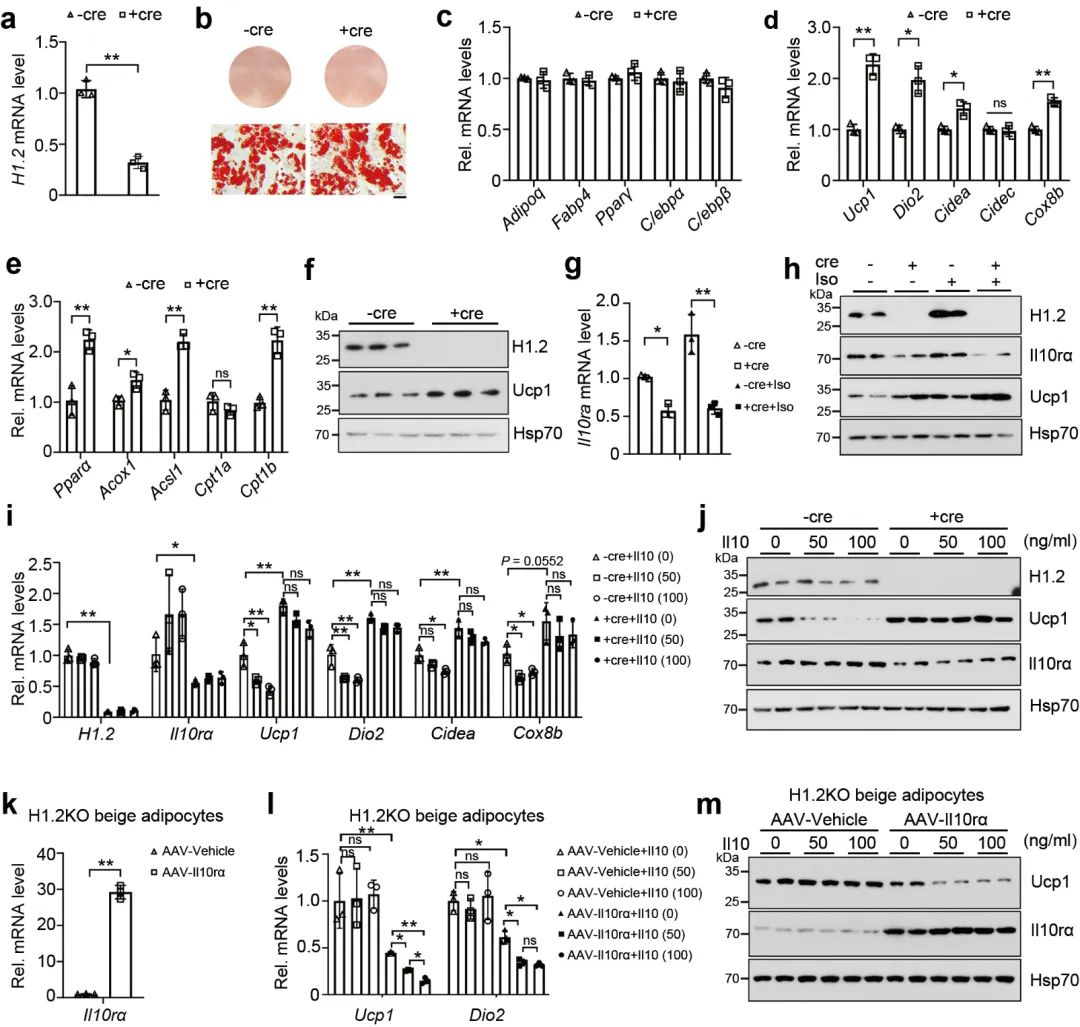
图 5 H1.2在体外调节米色脂肪细胞的Il10rα抑制产热
05.H1.2通过调节IL10rα影响冷诱导米色脂肪组织褐变
为了研究下调的IL10rα是否与冷刺激下H1.2AKO褐变有关, 在WT和H1.2 AKO小鼠的iWAT (AAV-Il10rα) 过表达Il10rα, 然后进行冷刺激, 通过qPCR, HE和免疫组化分析表明IL10rα的过表达减少了WT和H1.2AKO小鼠的iWAT的冷诱导褐变 (图 6a-g)。这些数据表明H1.2通过IL10rα抑制冷诱导的米色脂肪细胞产热。
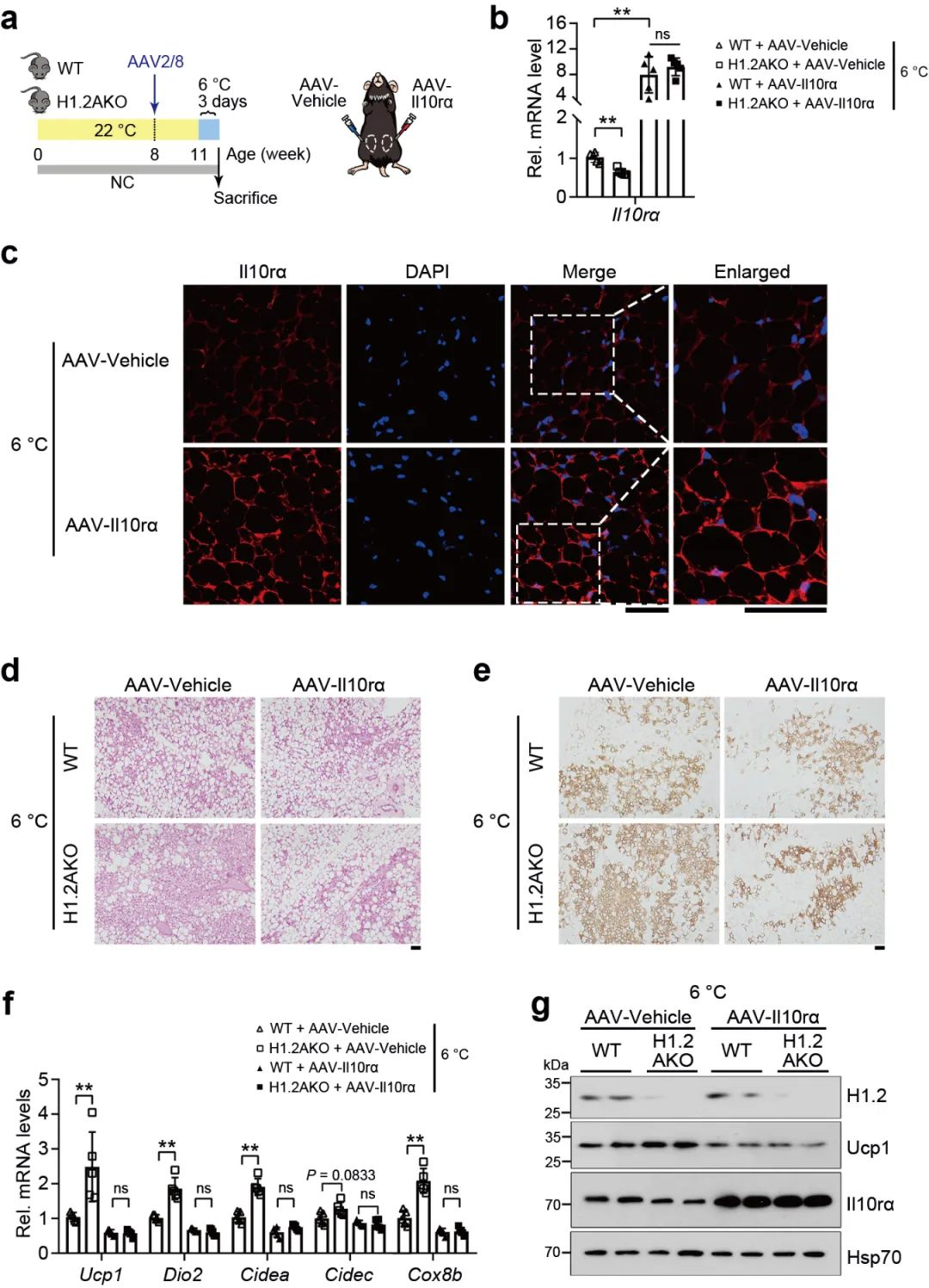
图 6 H1.2通过IL10rα调控冷诱导iWAT褐变
06.H1.2AKO小鼠能够通过IL10rα预防肥胖, 维持长期正常饮食状态下的能量稳态, 通过适应性产热减少体重增加
之前的研究表明, 米色脂肪细胞中H1.2参与产热, 因此研究者想要探究H1.2 AKO小鼠是否能够减轻脂肪堆积。研究者将H1.2 AKO和WT小鼠进行长期低脂饲养。结果表明, H1.2 AKO和WT小鼠的体重在刚开始没有明显差别, 但是18周龄时开始出现体重差异, 在28周龄时变得更加明显, H1.2AKO小鼠的脂肪质量和瘦肉质量增加较少 (图 7a-b)。另外, 28周龄的H1.2AKO小鼠的BAT和eWAT重量显著减少, 同时eWAT和iWAT的细胞大小变小 (图 7c-e)。在能量代谢方面, 28周龄的H1.2AKO小鼠, 氧气消耗和能量消耗增加, 葡萄糖耐量得到改善。同时, 在H1.2AKO小鼠的iWAT中, 脂肪生成基因下调, 产热基因和脂质β-氧化基因上调。这些结果表明H1.2AKO小鼠在长期低脂饲养后能够维持较好的代谢状态。
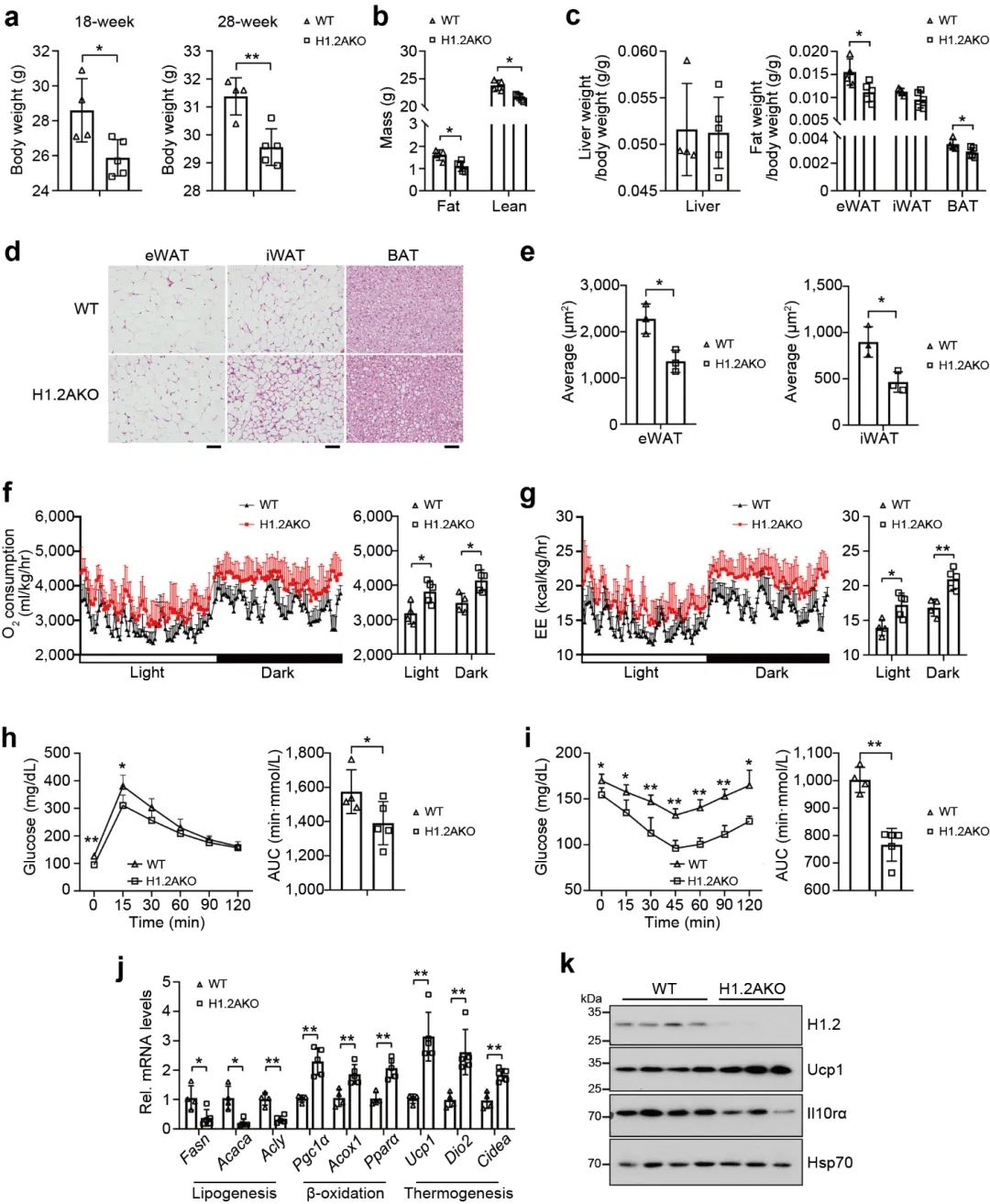
图7 H1.2AKO小鼠在长期正常饮食下具有较好的代谢状态
最近的研究表明, 与热中性条件 (30°C) 相比, 正常条件 (22°C) 对小鼠具有轻微的冷刺激。为了研究adrenaline信号诱导的适应性产热是否有助于长期维持低脂喂养的H1.2AKO小鼠的代谢表型, 将22℃饲养4周的小鼠置于30°C环境4个月, 以尽量减少适应性产热, 发现H1.2AKO小鼠和WT小鼠的体重, iWAT, eWAT和肝脏重量没有显著差别, 但是其BAT重量显著降低 (图8b-e)。同样, 与脂肪生成, 产热和脂质β-氧化相关基因的转录水平没有显著差别 (图8f, g)。说明脂肪细胞H1.2缺乏通过适应性产热减少体重增加。
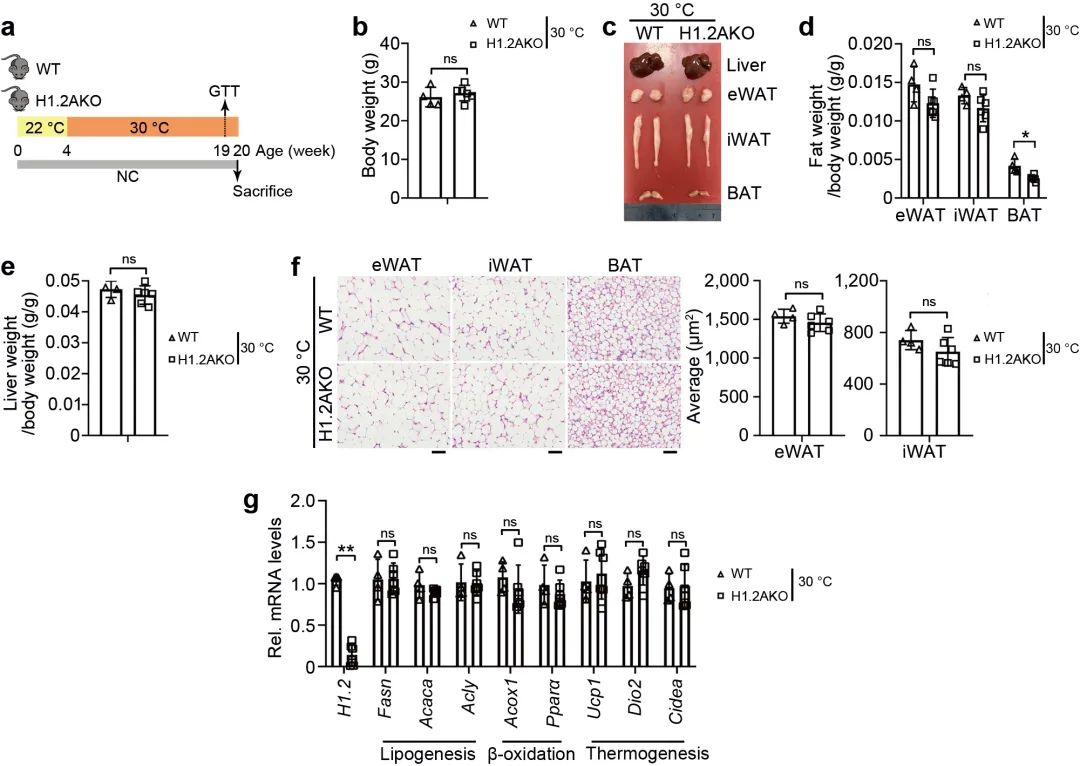
图8 脂肪细胞H1.2缺乏通过适应性产热减少体重增加
随后, 研究了高脂饲料喂养是否会影响热源性脂肪组织中H1.2的表达, 高脂饲养6个月后, 与低脂饲料喂养小鼠相比, iWAT和BAT中H1.2 mRNA和蛋白水平均升高 (图 9a-c); 同样, 不同体重指数的人体受试者的皮下脂肪中的H1.2基因和蛋白水平上调 (图9d)。回归分析表明BMI与皮下脂肪中 H1.2 mRNA水平呈正相关 (图 9e); 另外, H1.2AKO小鼠能够明显的缓解长期高脂饲养导致的体重, 脂肪重量的增加, 并且能够显著降低脂肪细胞大小, 增加能量消耗, 改善葡萄糖耐量, 增强胰岛素敏感性 (图 9d-k)。说明脂肪细胞中H1.2的缺失可以预防高脂饮食诱导的肥胖。
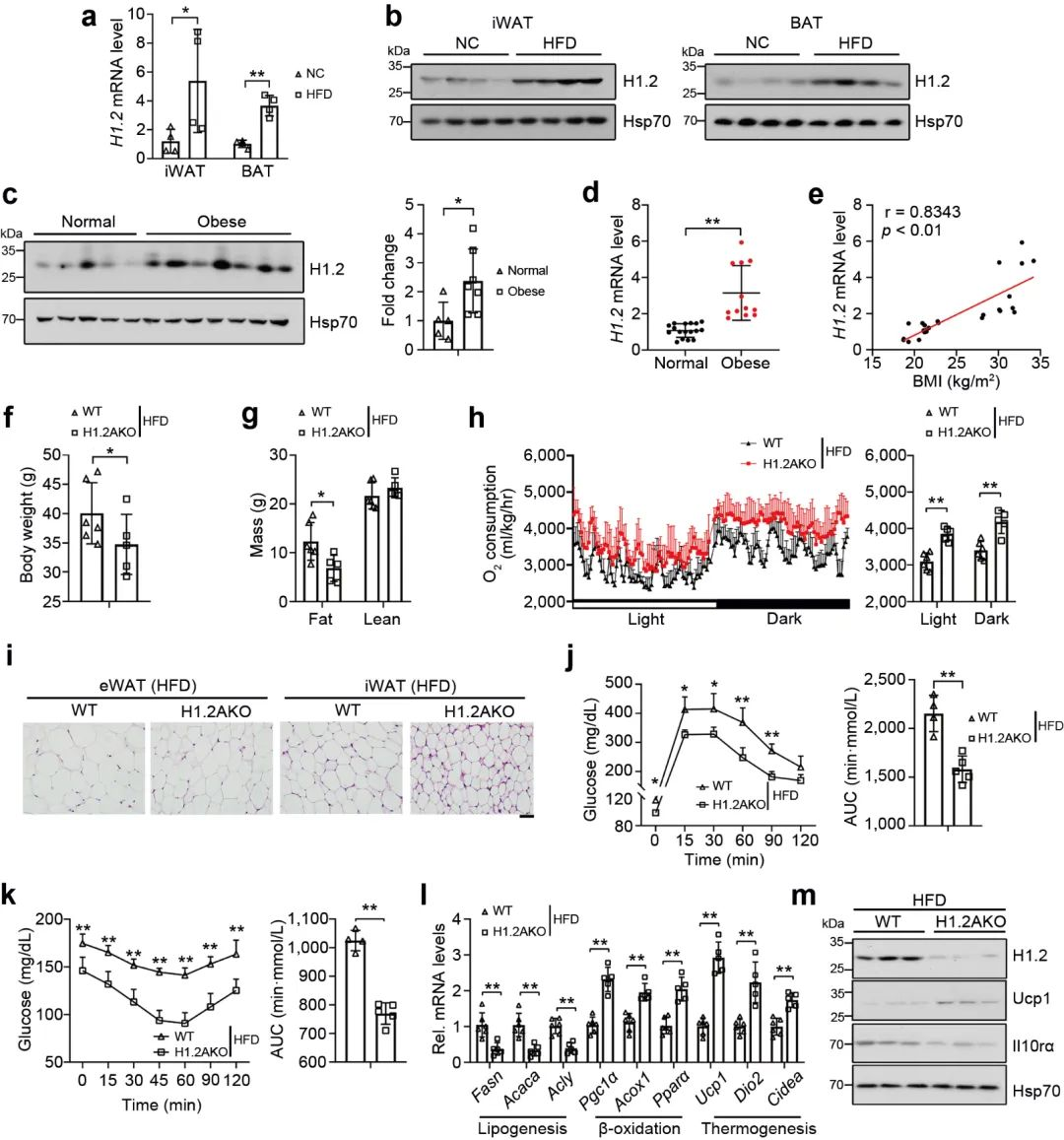
图9 H1.2AKO小鼠对高脂饮食诱导的肥胖具有抵抗作用
接下来, 研究者评估了H1.2是否依赖于IL10rα 抑制脂肪堆积, H1.2AKO和WT小鼠在低脂饲养10周后, 在iWAT中注射AAV-IL10rα, 再继续低脂饲养15周, 结果表明, Il10rα过表达可减弱脂肪细胞H1.2缺失对小鼠体重, 脂肪重量和各类脂肪组织的抑制作用 (图 10a-e); 同样, 过表达IL10rα后, H1.2AKO小鼠对葡萄糖耐量改善和胰岛素敏感性增强作用也被消除 (图 10g-h), qPCR结果表明产热基因和脂质β-氧化基因的上调也被抑制, 耗氧量率和能量消耗也降低 (图 10i-j)。这些结果说明H1.2AKO小鼠通过IL10rα预防肥胖。
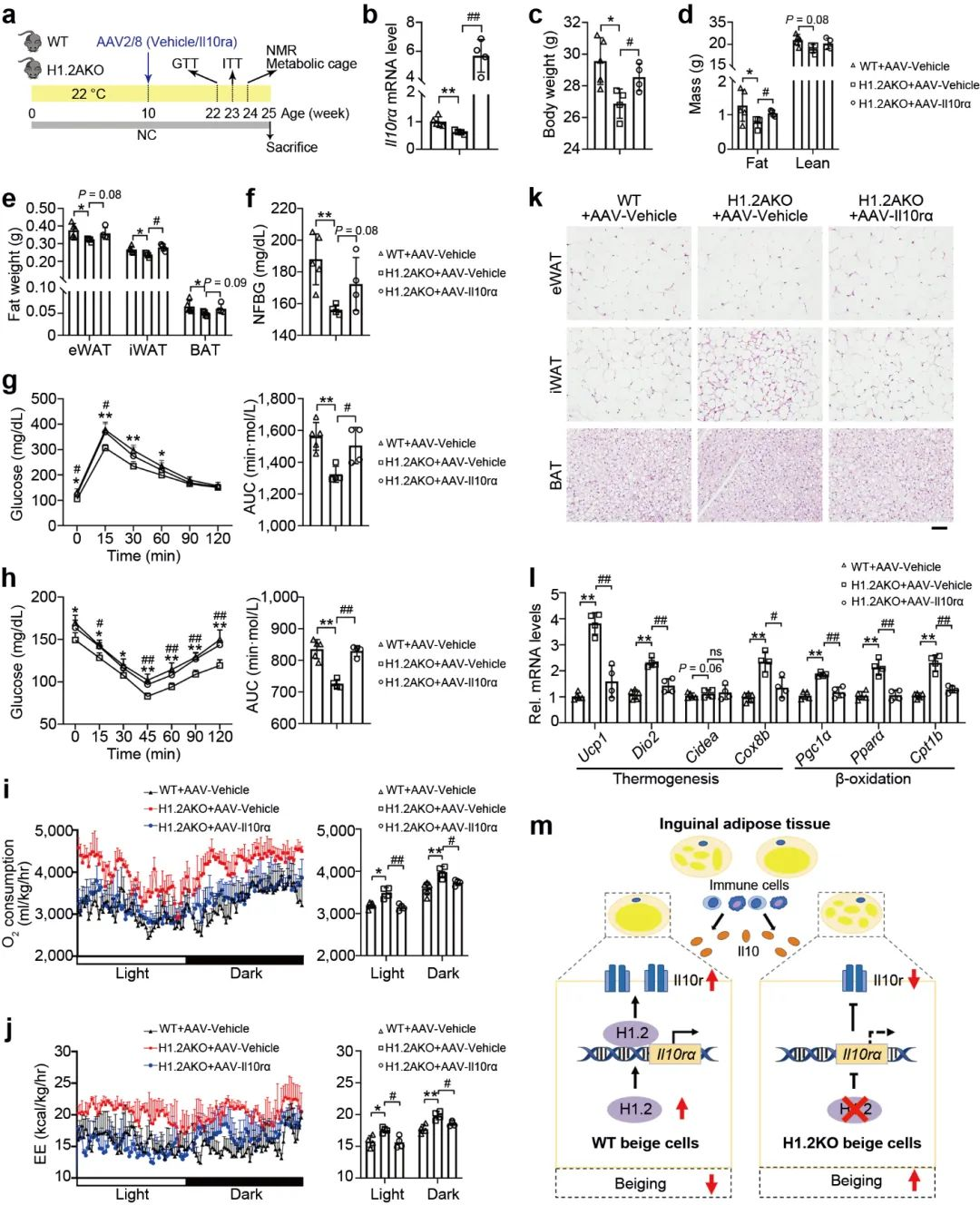
图10 IL10rα的过表达消除了H1.2AKO小鼠的代谢缓解作用
文章结论与展望
和元助力
和元生物有幸提供实验中使用的病毒包装(AAV-H1.2/AAV-IL10rα/Ad-Cre-H1.2)服务
业务咨询
更多活动详情可咨询(微信/电话):15800353038